Of chief importance in understanding the organization of the brainstem is knowledge of what is localized in each embryological subdivision and in any transverse section. This is a significant challenge for every student of neuroanatomy and it is to this challenge that we now turn in earnest. You have already faced the first step toward mastery of the essential knowledge: recognition of the external features of each brainstem subdivision, including the associated cranial nerves. After working through this chapter, you should be able to recognize how the numerous bumps and furrows along the length of the brainstem and spinal cord relate to the organization of internal structures that account for these external features. Before proceeding, it will be worth reminding yourself of the basic layout of sensory and motor neurons in the brainstem and spinal cord.
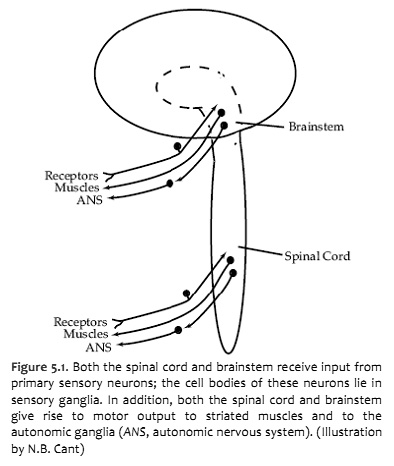
The central nervous system interacts with the outside world through primary sensory neurons, which convey information from the body or its environment into the brain and spinal cord, and motor neurons, which activate striated muscles and modulate the activity of cardiac and smooth muscles and glands (Figure 5.1). The cell bodies of primary sensory neurons lie in the dorsal root ganglia or the cranial nerve ganglia. Each neuron gives rise to a peripheral process, which receives information either directly or through association with receptors, and a central process, which enters the central nervous system and forms synapses with second order neurons. The cell bodies of somatic motor neurons lie in clusters or nuclei within the central nervous system and give rise to axons that innervate striated muscles in the body or head. In this chapter, you will be especially concerned with the organization of these second-order sensory neurons and somatic motor neurons. You will also be introduced to other motor neurons that are part of the visceral motor system (a.k.a., autonomic nervous system) and are indirectly responsible for governing cardiac muscle, smooth muscle or glands. So by the conclusion of this laboratory experience, you will learn how to locate the following nuclei:
nuclei that are the destination of all of the sensory input into the central nervous system (i.e., the location of all of the second-order neurons that receive the primary sensory input). There are two exceptions: the olfactory nerve and the optic nerve are not included in this discussion; for several reasons they are atypical nerves.
nuclei that are the origin of all of the output of the central nervous system (i.e., the location of all of the motor neurons and preganglionic visceral motor neurons).
From the viewpoint of neurology, the most important general principle of organization in the central nervous system is that each CNS function (e.g., perception of sensory stimuli, control of motor behavior) involves groups of neurons—interconnected through synapses—that are spatially distributed throughout several subdivisions. Groups of neurons that together subserve a particular function are called a ‘system’; for example, there are the visual, motor, and somatic sensory systems. The structures containing the neurons and axons of a particular system are collectively referred to as a ‘pathway’. (The term ‘system’ has a functional connotation, whereas the term ‘pathway’ refers to the structures involved.) Two important pathways are illustrated in Figure 5.2.
Note that receptors and muscles on the right side of the body are represented in the left side of the forebrain—a fact will be emphasized repeatedly over the next three weeks. Knowledge of the crossing of sensory and motor pathways as they ascend or descend provides critical information to the clinician who is localizing neurological lesions.
If damage to the central nervous system at every level gave rise to exactly the same signs and symptoms, it would not be worthwhile for you to learn the details of neuroanatomy that are presented in this Laboratory Guide. However, as neurologists and neuroscientists recognized long ago, the neurons involved in specific functions occupy specific locations in the central nervous system. Even those systems that are represented in multiple subdivisions bear different physical relationships to one another from one subdivision to the next (e.g., Figure 5.2). Because neurons that subserve specific functions occupy specific locations, the combinations of neurological signs and symptoms exhibited by particular patients often provide detailed information about the location of damage in the central nervous system.
2. Internal anatomy of the spinal cord
The following brief discussion of the internal anatomy of the spinal cord will introduce some of the general principles of organization that also hold true for the brainstem. A cross-section through the spinal cord is illustrated schematically in Figure 5.3. The gray matter forms the interior of the spinal cord; it is surrounded on all sides by the white matter. The white matter is subdivided into dorsal (or posterior), lateral, and ventral (or anterior) columns. Each of these columns contains bundles of axons related to specific functions. For example, the lateral columns are made up partly of axons that travel from the cerebral cortex to form synapses with motor neurons in the ventral horn. The dorsal columns carry much of the ascending sensory information from mechanoreceptors (much more on these long pathways in other course sessions).
The gray matter of the spinal cord is divided into dorsal and ventral (or posterior and anterior) ‘horns.’ The dorsal horn is the part of the gray matter that receives sensory information entering the spinal cord via the dorsal roots of the spinal nerves. (Not all sensory fibers terminate in the dorsal horn of the spinal cord; axons carrying sensory information from mechanoreceptors travel to the medulla before making their first synapse in the pathway to conscious perception; they will be covered later.) The ventral horn contains the cell bodies of motor neurons that send their axons out via the ventral roots to terminate on striated muscles. Thus, one important general rule of organization is that neurons in the spinal cord that process sensory information are spatially separate from motor neurons. (See Figure A6 and Table A1 of Neuroscience, 5th Ed., for more detail on the internal organization of spinal gray matter.)
As seen in the previous lab session, the inputs and outputs of the spinal cord are arranged segmentally into the 31 spinal nerves (Figure 5.4). However, the gray matter of the spinal cord is not obviously segmented. It can be thought of as a continuous column of cells running the length of the cord, although there are differences in the size of the dorsal and ventral horns at different levels. The dorsal and ventral horns are largest where they supply the upper and lower limbs because there are significantly greater numbers of outgoing and incoming nerve fibers at those levels. There is also variation along the length of the cord in the number of fibers in the columns of white matter (and, therefore, in their relative size). The amount of white matter is greatest at cervical levels and least at sacral levels. This is because ascending and descending fibers from and to all levels must pass through the cervical cord.
Challenge 5.1—internal anatomy of the spinal cord
With reference to Figures 5.4 and 5.5 and the chart below, carefully inspect the internal features of the spinal cord that are present in each segment, as well as those that are different (or present in only in one segment). To complete this challenge, spend some time browsing the spinal cord sections inSylvius4, and find each of the internal features identified across the upper row in the chart below in theSylvius4spinal cord.
Internal features
Spinal cord segment
Dorsal horn
Lateral horn
Ventral horn
White matter
Gracile tract
Cuneate tract
Lateral corticospinal tract
Ventral corticospinal tract
Anterolateral system
Cervical segments (8)
--
+++++
Thoracic segments (12)
++++
Lumbar segments (5)
--
+++
--
Sacral segments (5)
--
++
--
Coccygeal segment (1)
--
+
--
--
3. The internal anatomy of the brainstem
The internal organization of the brainstem is considerably more complicated than that of the spinal cord. However, two factors work in your favor as you study its features. First, important general principles of organization of the spinal cord also hold true for the brainstem. Second, much of the complexity of the brainstem is contributed by cell groups and axon tracts that will not be considered in this course. In the following discussion, the general plan of organization of the brainstem is presented first. Next, the prominent internal features that characterize each subdivision are identified. Finally, the cranial nerve nuclei are discussed in some detail. An understanding of their functions and locations is essential for diagnosing (and treating) neurological disease.
It would be convenient if each subdivision of the brainstem were sufficiently homogeneous along its length that one cross-section could serve as a ‘typical’ representative for the entire subdivision. However, the brainstem changes continuously along its length—the subdivision into three parts is somewhat arbitrary. As a compromise between examining three sections (one for each subdivision) and hundreds, seven sections of the brainstem are shown to serve as representatives (Figure 5.6).
Once you understand the organization of these seven levels and the way various pathways traverse them, you should be able to identify the location of any section through the brainstem and the important pathways represented in it.
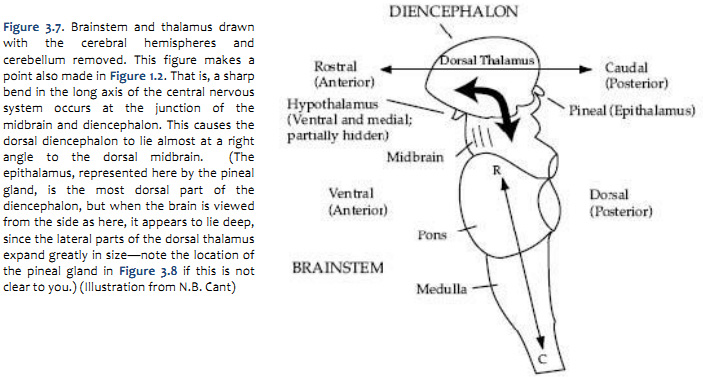
A schematic overview of the levels of the brainstem to be discussed is presented in Figure 5.7. At this stage, it is not important to study the details; we will come back to them. For now, three points should be taken from the figure:
All of the sections are shown at the same magnification. In most atlases (including Sylvius4), the smaller sections are magnified more than the larger ones, and it is easy to lose sight of the relative proportions of the different subdivisions.
The cranial nerve nuclei lie in the tegmentum of the brainstem, as do many of the major ascending and descending tracts.
Just as in the spinal cord, the nuclei that receive sensory inputs via the cranial nerves are spatially separate from those that give rise to motor output. The sensory nuclei are located laterally in the brainstem, whereas the motor nuclei are located medially. The spatial segregation of sensory and motor functions provides an important clue for localization of focal damage in the brainstem.
The Brainstem Cross Sectional Atlas in Sylvius4 contains 14 sections through the brainstem and four spinal cord sections featuring segmentation of well over 100 neuroanatomical structures. This atlas should be used to fill-in the gaps between the seven sections illustrated in this chapter.
As you work through these chapter figures and the associated legends, find the same structures one at a time in the Brainstem Cross Sectional Atlas in Sylvius4. The sections shown here are actually re-colorized versions of the same sections that are in Sylvius4. As suggested in Lab 1, you may wish to look-up each labeled term (and each term that appears in bold in the figure legends) in Sylvius4’s Visual Glossary module; there, you can ‘bookmark’ each term for further study and review.
Since we are not yet studying the functional significance of many of these structures (we will do so over the next two weeks), you should click on the structures identified in the figures, and when you do so, that structure becomes selected and a brief overview of that structure becomes available in the text window.
Tip—click on the loudspeaker icon at the bottom of the text window to hear the proper pronunciation of any term that is unfamiliar to you.
Tip—to increase the magnification of the image, grab the lower-right corner of the image window and pull the window to its maximum extent.
Challenge 5.2—internal anatomy of the medulla
With reference to Figures 5.8 & 5.9 and the chart below, carefully inspect the internal features of the medulla from its caudal union with the spinal cord to the pons. Spend some time browsing the six medullary sections in Sylvius4, and find each of the internal features described in the chart below.
pair of extended longitudinal bulges on either side of a deep midline furrow; technically, this bulge is called the tuberculum gracilis, which is formed by the underlying gracile tract
continuation of the tract of the dorsal spinal cord
Gracile tract & nucleus
medial, superficial bundle of myelinated axons arising from the dorsal column of the spinal cord
just deep to the gracile tract is the gracile nucleus, a compact gray matter structure that receives the synapses made by gracile tract axons
Cuneate tract
(dorsal surface)
pair of extended longitudinal bulges just lateral to the gracile tracts; technically, this bulge is called the tuberculum cuneatus, which is formed by the underlying cuneate tract
continuation of the tract of the dorsal spinal cord
Cuneate tract & nucleus
just lateral to the gracile tract, superficial bundle of myelinated axons arising from the dorsal column of the spinal cord
at the superior “head” of the cuneate tract is the cuneate nucleus, a compact gray matter structure that receives the synapses made by cuneate tract axons
Pyramidal decussation
(ventral surface)
see Medullary pyramids below
apparent “stitching” of fibers that cross the midline
Pyramidal decussation
see Medullary pyramids below
midline crossing of dense bundles of myelinated axons that run the longitudinal extent of the ventral brainstem
accounts for the formation of the lateral and ventral (anterior) corticospinal tracts of the spinal cord
pair of extended longitudinal bulges on either side of a deep midline furrow
Medullary pyramids
dense bundle of myelinated axons that run the longitudinal extent of the ventral brainstem; these axons are also known as the corticospinal tract
these same axons are present in the internal capsule, cerebral peduncles, basilar pons, and about 90% are present in the lateral columns of the spinal cord
Inferior olive
(ventral-lateral surface)
pair of elongated bulges just lateral to the pyramids; a shallow furrow separates the pyramid and olive on each side
Inferior olivary nucleus
prominent nucleus of the ventral-lateral medulla just dorsal to the medullary pyramids
note the highly convoluted bands of gray matter that account for the superficial, ventral-lateral bulge
Hypoglossal nerve (XII)
(ventral-lateral surface)
exits through ventral-medial surface
Hypoglossal nerve roots & nucleus
nerve roots emerge between the medullary pyramid and the olive
trace these nerve roots dorsally to their origin in the hypoglossal nucleus, located along the dorsal midline
Challenge 5.3—internal anatomy of the pons
With reference to Figures 5.10 & 5.11 and the chart on the following page, carefully inspect the internal features of the pons. As you did for the medulla, spend some time browsing the sections inSylvius4, and find each of the internal features described in the chart below.
massive system of transverse fibers that “bridge” the longitudinal axis of the brainstem; these fibers originate in the basal region of the pons and continue around its ventral-lateral aspect to enter the cerebellum
the ventral half of the pons (also called the basilar pons) contains gray matter, longitudinal axons, and transverse fibers called the pontocerebellar fibers that decussate and form the contralateral middle cerebellar peduncle
these fibers arise from a scattering of gray matter in the basilar pons, called the pontine nuclei, and terminate in the contralateral cerebellum
also in the basilar pons are prominent fascicles of axons from the cerebral cortex that project to various nuclei of the brainstem and the spinal cord; collectively, these are the corticobulbar/corticospinal fibers
Trigeminal nerve (V)
(ventral-lateral surface)
enters/exits pons by penetrating the transverse, pontocerebellar fibers
Trigeminal nerve roots & nucleus
trace the nerve V roots dorsally to their origin in the trigeminal nuclear complex; at this level, note the location of the trigeminal motor nucleus and, just lateral to it, the principal (chief sensory) nucleus
now, keep your eye in this same general region and section caudally: in this same dorsal-lateral position in the caudal pons and throughout the medulla, the spinal trigeminal nucleus and the spinal trigeminal tract are present (the spinal nucleus can be further subdivided)
enters/exits near the midline at the pontomedullary junction (most medial of the three that emerge from this junction)
Abducens nerve roots & nucleus
explore the medial tegmentum of the pons and locate nerve VI roots; note how they course through the basilar pons just lateral to the corticobulbar/corticospinal fibers
trace these nerve roots dorsally to their origin in the abducens nucleus, which is located along the dorsal midline
Facial nerve (VII)
(ventral-lateral surface)
enters/exits through ventral-lateral surface at pontomedullary junction (middle of the three that emerge from this junction)
Facial nerve roots & nucleus
explore the lateral tegmentum of the pons and locate nerve VII roots; note how they trace a most unusual trajectory around the dorsal aspect of the abducens nucleus (cf. Figure 5.14)
it may not be possible to trace these nerve roots all the way back to their origin in the facial nucleus, which is located just medial and ventral to the trigeminal nuclear complex
nerve VII roots exit the facial nucleus medially, then course dorsally around the abducens nucleus, and finally ventral-laterally toward a lateral exit (this is how nerve VII ends up being lateral to nerve VI)
enters through ventral-lateral surface at pontomedullary junction (most lateral of the three that emerge from this junction)
Vestibular nuclear complex
explore the lateral tegmentum of the pons and locate nuclei of the vestibular nuclear complex (you may not find any nerve VIII roots in these cross sections); you will find the vestibular nuclei dorsal to the trigeminal nuclear complex and spinal trigeminal tract
find the superior, lateral and medial vestibular nuclei
section down into the medulla and locate the spinal vestibular nucleus
so what about the cochlear division of nerve VIII? It terminates in a superficial nucleus of the dorsal-lateral upper medulla called the cochlear nucleus. Although not labeled in Sylvius4, it is visible in the section labeled “8-Medulla” as the gray matter that wraps around the dorsal-lateral surface of the inferior cerebellar peduncle
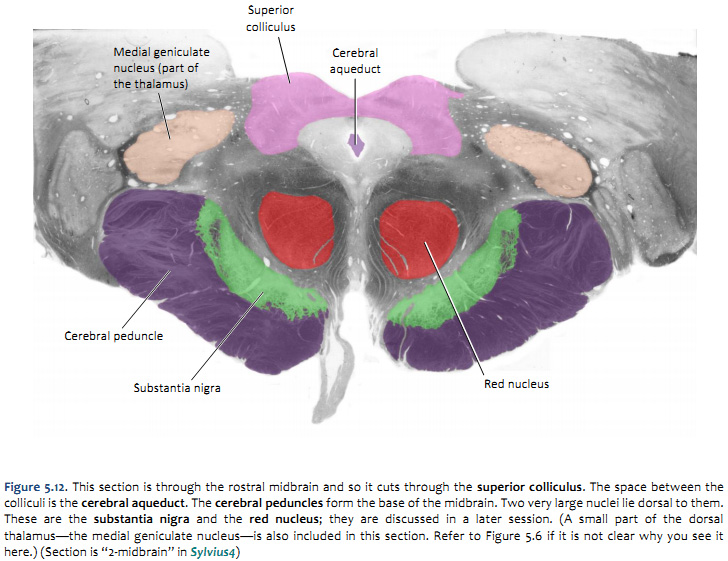
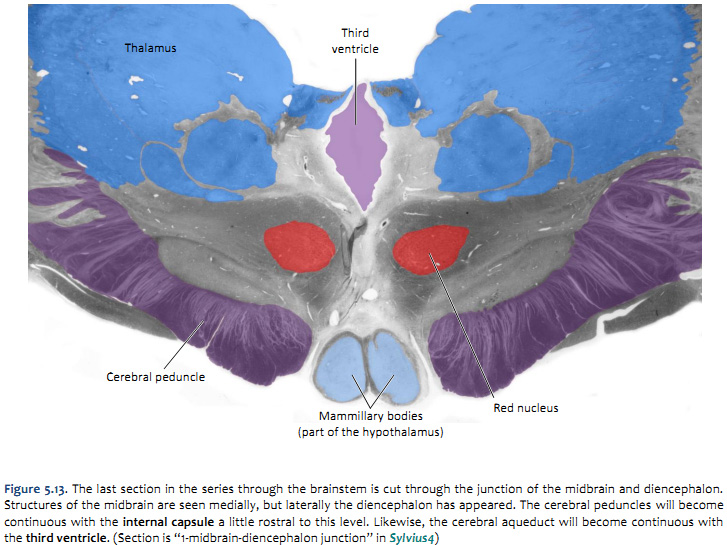
Challenge 5.4—internal anatomy of the midbrain
With reference to Figures 5.12 & 5.13 and the chart on the following page, carefully inspect the internal features of the midbrain. As you did for the medulla and pons, now spend some time browsing the sections inSylvius4, and find each of the internal features described in the chart below.
large, longitudinal “stalks” (peduncle mean stalk) that occupy the ventral midbrain
Cerebral peduncles
technically, “cerebral peduncle” refers to the entire ventral midbrain, including the midbrain tegmentum and the fiber systems that run through the stalks; however, it is common to use the term “cerebral peduncle” to refer specifically to these fiber systems (the proper term for these ventral portions of the peduncles—where the fibers are—is basis pedunculi)
the cerebral peduncles comprise efferent fibers of the cerebral cortex that terminate in the brainstem and spinal cord; these fibers are referred to collectively as the corticobulbar / corticospinal fibers; this compound terms indicates that the some of these fibers terminate among brainstem nuclei (“bulbar” refers to the brainstem and cranial nerve nuclei), while other fibers continue through the brainstem and terminate in the spinal cord
it is important to recognize the course of these fibers from their origin in the cerebral cortex through brainstem: cerebral cortex → subcortical white matter → internal capsule → cerebral peduncle → basilar pons → medullary pyramids → lateral and anterior (ventral) corticospinal tract
there are about 20 million axons in each cerebral peduncle; can you guess how many axons are present in medullary pyramid by simply noting the difference in size of these two structures?1 (the majority of these axons never reach the spinal cord)
now consider the tegmentum of the midbrain; just dorsal to the cerebral peduncles (basis pedunculi) there is an important gray matter nucleus called the substantia nigra, which you will learn much more about when we study the basal ganglia
and just dorsal to the substantia nigra, is a spherical gray matter structure called the red nucleus, which you will study when we consider cerebellar systems
Oculomotor nerve (III)
(ventral surface)
exits through ventral surface just medial to cerebral peduncles (in the interpenducular fossa)
Oculomotor nerve roots & nuclear complex
trace these nerve roots dorsally to their origin in the nuclei of the oculomotor complex along the midline of the dorsal tegmentum; here you will find two divisions: the oculomotor nucleus and the Edinger-Westphal nucleus
this nuclear complex is embedded within a large region of gray matter that surrounds the cerebral aqueduct, termed the periaqueductal (or central) gray
Inferior colliculi
(dorsal surface)
inferior pair of the four bumps that are visible in brainstem model/illustration, but are normally covered by the cerebellum
Inferior colliculi
in the caudal midbrain, the inferior colliculi are gray matter structures that occupy a position just dorsal and lateral to the periaqueductal gray (see section “3 - Midbrain”)
together with the superior colliculi, they form the “roof” of the midbrain (above the cerebral aqueduct); for this reason, these four bumps are also called the tectum (tectum means roof)
the trochlear nerve exits the dorsal surface of the brainstem just caudal to the inferior colliculus (see Brainstem Model in Surface Anatomy module)
although that nerve is not visible in section 3 – Midbrain, you can see the small trochlear nuclei where you should expect to find somatic motor nuclei, along the midline of the dorsal tegmentum
Superior colliculi
(dorsal surface)
superior pair of the four bumps that are visible in brainstem model/illustration, but are normally covered by cerebellum
Superior colliculi
in the rostral midbrain, the superior colliculi are laminated gray matter structures that occupy a position just dorsal and lateral to the periaqueductal gray matter (see section lableled “2 - Midbrain”)
together with the inferior colliculi, they form the “roof” (tectum) of the midbrain (above the cerebral aqueduct)
1There are about one million axons in each medullary pyramid.
4. The cranial nerve nuclei
The cranial nerve nuclei are made up of the neurons in the brainstem that receive primary sensory inputs or that give rise to motor outputs. Just as there are cell groups in the dorsal horn of the spinal cord that receive sensory information and cell groups in the ventral horn that contain motor neurons, there are separate sensory and motor nuclei in the brainstem.
The spinal cord receives sensory information from the body surface, muscles (via the muscle spindles) and the viscera, and sends motor axons to striated muscles and to the autonomic ganglia. These same inputs and outputs exist for the head. In addition, there are specialized sensory inputs in the head that do not have equivalents in the spinal cord. These include the inputs for hearing, balance and taste. The motor outputs are just a little more complicated as well. There are nuclei which innervate the extrinsic eye muscles and the tongue. Since the muscles that are innervated are derived from somites, these motor neurons are exactly equivalent to those in the ventral horn, which also innervate muscles derived from somites. In addition, there are nuclei in the brainstem that innervate the muscles derived from the branchiomeres of the pharyngeal arches (jaw muscles, muscles of facial expression, the pharynx and larynx). There is only one such cell group in the spinal cord; it innervates the trapezius and sternocleidomastoid muscles, which are also derived from branchiomeres. It is included with the cranial nerve nuclei, since it gives rise to the spinal part of cranial nerve XI. Finally, there are cell groups in the brainstem that form part of the visceral motor (autonomic) system and send axons to autonomic ganglia in organs throughout the body. By working through Challenges 5.2–5.4, you have already encountered many of these nuclei; let’s now build a stronger appreciation for the organization of these nuclei in each division of the brainstem.
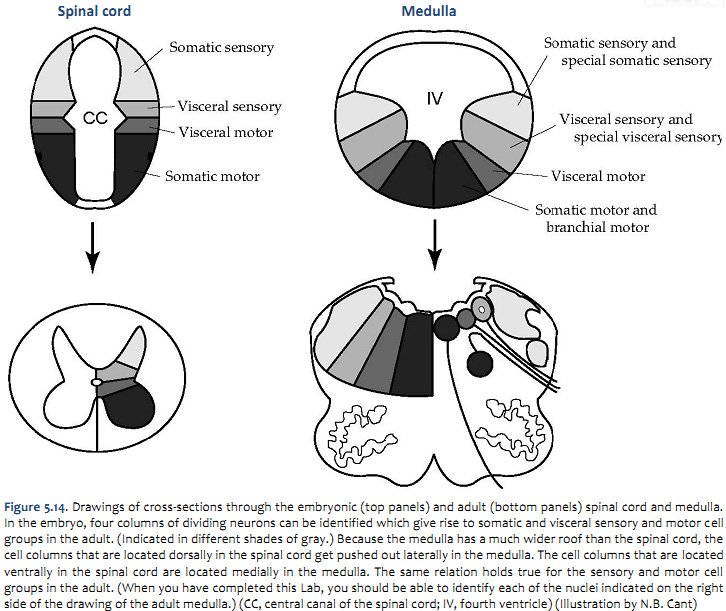
The organization of the sensory and motor neurons in the spinal cord and brainstem are similar, for reasons that are clear when one considers the development of these two regions of the neural tube—as explained in Figure 5.16. Sensory neurons, which are located dorsally in the spinal cord, are located laterally in the medulla and pons. Motor neurons, which are located ventrally in the spinal cord, are located medially in the medulla, pons, and midbrain.
Ten sets of cranial nerves are associated with the brainstem. These nerves supply sensory inputs to or derive motor output from 16 sets of cranial nerve nuclei. In the following discussion, ways of grouping these nuclei to make them easier to remember are presented. Next, the way in which they match up with the nerves will be described. Finally, they will be located on the cross-sections of the brainstem with which you are already familiar.
One further point can be made from Figure 5.14. In the medulla, the motor column that gives rise to somatic motor nuclei also gives rise to the motor neurons that will innervate the branchiomeric muscles. These motor neurons migrate from the midline into the ventrolateral part of the tegmentum and send their axons out in nerves that exit the brain laterally (i.e., V, VII, IX, X, and XI). The somatic motor neurons, on the other hand, send their axons out ventrally (in a line with ventral roots of the spinal cord), except for the trochlear nucleus as noted above. Oddly, the parasympathetic preganglionic neurons (visceral motor neurons) in the medulla and pons also send their axons out laterally; in the spinal cord (and in the midbrain), the autonomic preganglionic axons leave in the ventral roots. All of the sensory inputs enter the brain laterally.
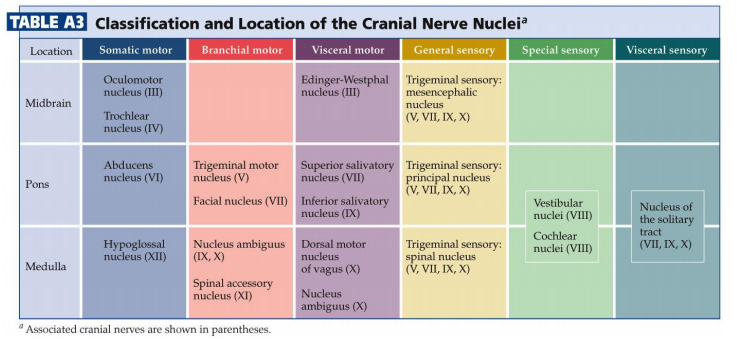
The sixteen cranial nerve nuclei can be most easily remembered if they are assembled into functional groups and anatomical location (Table A3—from Purves et al., Neuroscience, 5th Ed.; Figure 5.15). Three of the groups are motor nuclei (a somatic motor group, a branchial motor group, and a parasympathetic preganglionic group). The other three groups are sensory nuclei (general sensory, special sensory, and visceral sensory).
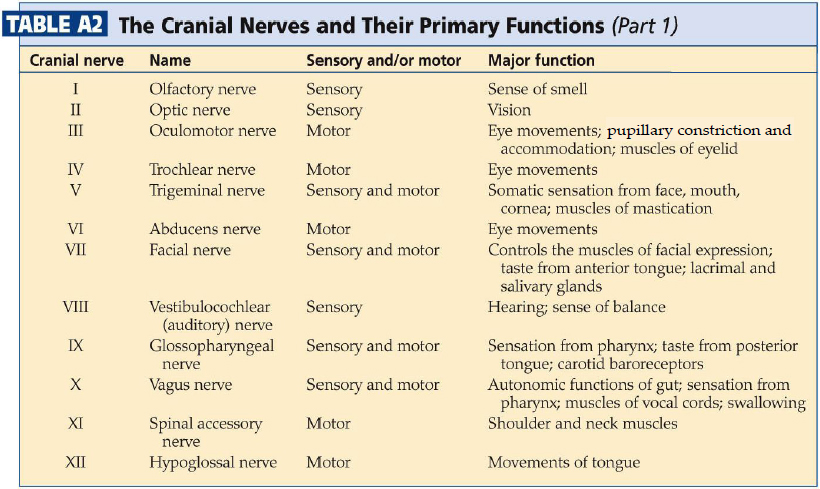
How do these nuclei relate to the components of the cranial nerves that you studied in Lab 4? Table A2lists the cranial nerve nuclei from which the sensory and motor components of each nerve arise.
Table A2 illustrates the point that most of the cranial nerves are connected to only one or two cranial nerve nuclei. Only three nerves carry components from more than two nuclei (ignore nerve XI in this regard). These nerves—VII, IX, and X—each carry five components: a branchial motor component, a parasympathetic component, a somatic sensory component, a special visceral sensory component (taste), and a general visceral sensory component. These three nerves lack a somatic motor component.
5. Location of the cranial nerve nuclei in brainstem cross-sections
Some of the cross-sections which were introduced earlier are reproduced in Figures 5.16, 5.17, 5.18, and 5.19. The locations of most of the cranial nerve nuclei listed in Tables A2, &A3 are indicated on these sections.
Only four of the cranial nerve nuclei are not identified in the preceding figures. The spinal accessory nucleus is found in the first few cervical segments of the spinal cord. Very few people in the world know just where the superior and inferior salivatory nuclei are located (somewhere in the dorsal tegmentum of the rostral medulla/caudal pons). The cochlear nucleus is located just at the junction of the medulla and pons (in the same general location as the vestibular nucleus).
You should now be able to easily identify the brainstem subdivisions from which each section is taken. You should also be able to identify each of the cranial nerve nuclei (except for the four mentioned in the preceding paragraph). To consolidate your understanding of the surface anatomy of the brainstem and of sections through it, identify the point at which each cranial nerve enters or leaves the brain, and determine the approximate trajectory of the axons as they travel to or from this point to the cranial nerve nuclei with which they are connected. By the end of this laboratory experience, you should be able to view a cross section through the brainstem and identify the landmarks that characterize the subdivision, the level of the brainstem from which it was taken and the locations of the cranial nerves and nuclei that are present. Make sure that you can identify all of the structures (in bold) discussed in this chapter and identified in the figures.
6. Signs and symptoms associated with specific lesions of the brainstem
Figure 5.20 contains examples of four different lesions that are commonly seen after vascular accidents involving the brainstem. Consider each of these sections in turn, and work through the final challenge of this lengthy chapter: predicting the functional impairments that result from stroke in the small circumferential or penetrating vessels that supply the brainstem.
Challenge 5.5—predicting functional impairment post stroke
Selective occlusion or hemorrhage of specific arteries and arterioles that supply the brainstem can lead to damage that is preferentially localized to its medial or lateral parts. This is because, as detailed in Lab 2, the vascular supply to the medial brainstem is distinct from that to the lateral brainstem. Using the information in Labs 2 & 4, as well as the sections in Sylvius4, identify which cranial nerve structures are likely to be damaged in each of the lesions indicated in Figure 5.20.