|
|
Lab 2 - Functional Organization of the Cerebral Cortex and Blood Supply |
||
| Reading: | ||
| Coursera Media: | Lab 2 Overview
Now that you have been introduced to the landmarks of each cerebral lobe, let’s look more closely at the complex patterns of gyri and sulci that are important to recognize. The reason for doing so is to establish a common language (a nomenclature) and a robust mental model of cerebral hemispheres that we will use as we study and learn the functions of different regions of the cerebral cortex, as well as how they interact in networks that span multiple lobes of the cerebral mantle—and even the two hemispheres. So let’s begin with the lateral aspect of the human brain and the frontal lobe.
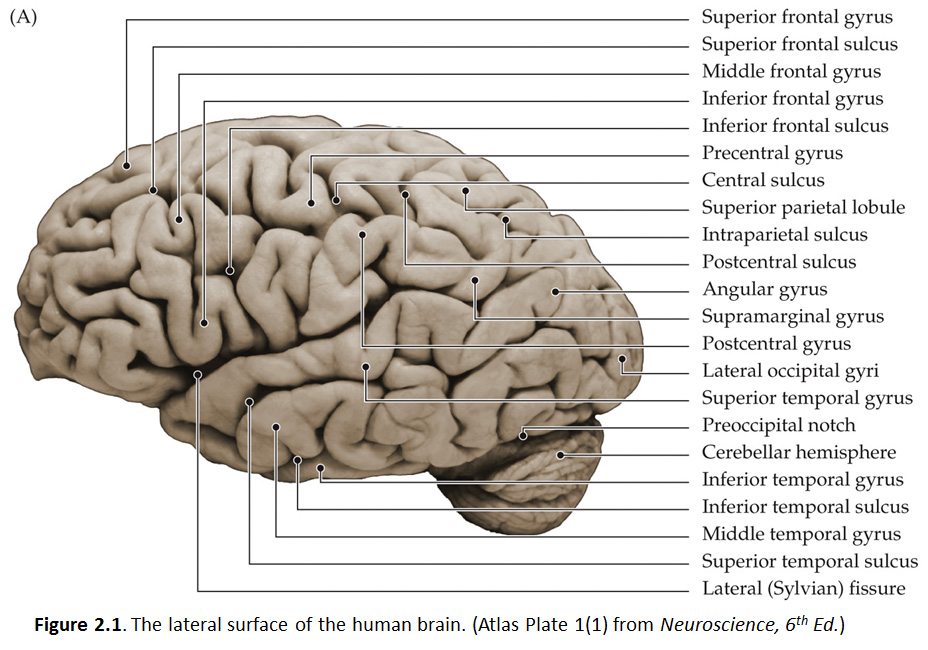
As you now know, the frontal lobe is the most anterior of the four lobes and is separated from the parietal lobe by the central sulcus (Figure 2.1; see also Grants 7.86A and Netters 99A); it is certainly one of the most important landmarks in the cerebral cortex. As you will study in detail elsewhere in this course, neurons in deeper layers of the precentral gyrus grow axons to the motor nuclei of the brainstem and spinal cord that, in turn, innervate the striated muscles of the body. Fine, skilled movements are dependent on the integrity of the motor cortex and, of course, the axons extending from it. The axons that arise from neurons in the motor cortex and extend to the spinal cord are known as the corticospinal tract. Those axons that extend from the motor cortex to motor nuclei in the brainstem are known as the corticobulbar tract. (The brainstem is sometimes referred to as ‘bulbar’ because it has a shape resembling a bulb.) While you can’t see the individual fibers that make up the tracts, you can see the structures through which they pass as they course between the cortex and their targets in the brainstem and spinal cord (we will identify them later in the course). The gyral formations anterior to the precentral gyrus on the dorsal-lateral aspect of the frontal lobe between the lateral fissure and the longitudinal fissure can be recognized as three parallel, longitudinal gyri (Figure 2.1). Adjacent to the longitudinal fissure is the superior frontal gyrus, which is typically the most obvious of the three. This gyrus continues on the medial surface of the hemisphere in the depths of the longitudinal fissure; we will see it again when we examine more closely the midsagittal plane. Just inferior to the superior frontal gyrus is the middle frontal gyrus, with the superior frontal sulcus separating the two. Note that these two gyral formations are fairly straight and parallel to the longitudinal fissure in the parasagittal plane. At their posterior end, they sometimes merge with the precentral gyrus. However, there is often an interrupted (discontinuous) sulcus that separates them from the precentral gyrus, named the precentral sulcus. Just inferior to the middle frontal gyrus, across the inferior frontal sulcus, is a much more complex gyral formation called the inferior frontal gyrus. For our purposes, we won’t be concerned with the names of the subcomponents of this complex gyrus or their differential functional contributions to cognition and behavior. Suffice it to say that the inferior frontal gyrus contains a critical functional division of the motor cortex that participates in the production of speech and non-vocal forms of language expression. This division has a special name, Broca’s Area, given in honor of the famous French neurologist, Pierre Paul Broca, who first recognized the significance of this gyral formation for human speech in the mid-19th century. Interestingly, the left hemisphere is dominant in most individuals for this function (especially in males and right-handers), such that damage to the left inferior frontal gyrus is more likely to produce an impairment of language expression, called Broca’s aphasia (aphasia means “without speech”), than a comparable lesion involving the right inferior frontal gyrus. Interestingly, this is also a division of the premotor cortex where in non-human primates neurons with “mirror” properties have been characterized. That is, neurons in the posterior part of the inferior frontal gyrus fire when certain actions are executed and when those same actions are observed (mirrored) in the behavior of another individual. Whether these neurons participate in imitation learning—or perhaps even decoding the intentions of others and empathy—remains an area of intense investigation and debate. Next, let’s cross the lateral fissure. The gyral structure that forms the inferior bank of the lateral fissure is the superior temporal gyrus. This gyrus is arranged parallel to the lateral fissure and extends from the anterior pole of the temporal lobe to the parietal lobe (specifically, the angular gyrus of the inferior parietal lobule). The elongation of this structure in nearly the horizontal plane establishes a framework for considering the remaining temporal gyral formations of the lateral surface. Including the superior temporal gyrus, it should be possible to recognize three elongated gyral structures that lie parallel to the lateral fissure: the superior, middle and inferior temporal gyri. The superior and middle temporal gyri are separated by the superior temporal sulcus and the middle and inferior gyri are separated by the inferior temporal sulcus, although the latter sulcus is often discontinuous and sometimes difficult to recognize. Notice how the lateral aspects of the frontal and temporal lobes bear some resemblance (at least conceptually): they both comprise three, parallel longitudinal gyri that extend from the anterior pole of each lobe back toward the parietal lobe (and the occipital lobe in the case of the temporal gyri). Once you are familiar with these gyral features in one specimen, spend some time examining as many specimens as are conveniently available in the laboratory. Once again, look for differences and common structural themes among brains and between hemispheres. In particular, carefully examine the pattern and length of the lateral fissure and consider the following questions.
These questions have been asked and addressed in studies of hemispheric asymmetries and gender differences related to language lateralization in the human brain. The superior aspect of the temporal lobe contains the cortical divisions whose functions pertain to audition and the reception of language. The posterior aspect of the superior temporal gyrus is often (but not invariably) associated with a special functional name, Wernicke’s Area, named in recognition of the seminal contributions of the 19th century German physician, Carl Wernicke, who described a disturbance of understanding speech, now called Wernicke’s aphasia. You may find differences in the structure of the superior temporal gyrus in the two hemispheres reflected in the pattern and length of the lateral fissure; such differences may, in turn, be related to the language dominance of the left hemisphere. Hemispheric differences in this cortical region likely also pertain to lateralized auditory functions, and may even relate to musical abilities and other special talents that pertain to audition and semantic encoding. As you will learn elsewhere in the course, critical language-specific nodes in Wernicke’s area may be found just about anywhere on the lateral aspect of the temporal lobe in some people, and even into the inferior aspect of the lateral parietal lobe (inferior parietal lobule). Other regions of the temporal lobe serve visual processing (inferior aspect), emotional processing (anterior-medial), memory and spatial navigation (posterior-medial), and the integration of sensory experience (posterior-lateral); we will consider these functions later in the course. Let’s return to the superior side of the lateral fissure. Since you already worked through recognition of the central sulcus, you should now be able to view the lateral surface of any hemisphere and know to look for the central sulcus lazily coursing from the longitudinal fissure over the dorsal-lateral surface of the cerebrum in a lateral-ventral and slightly anterior direction (hopefully, the directional terms applied to the forebrain are also becoming second-nature to you). Much of the cortex that you observe posterior to the central sulcus is part of the parietal lobe. The prominent gyral structure that forms the posterior bank of the central sulcus is the postcentral gyrus, which is mostly parallel to the precentral gyrus. This gyrus is concerned with somatic sensation and will be a major focus for clinicians concerned with the cerebral localization of touch and body position sensations. Immediately posterior to the postcentral gyrus is the postcentral sulcus, which separates the postcentral gyrus from two major gyral formations of the parietal lobe: the superior and inferior parietal lobules. The boundary between these two lobules is often difficult to appreciate; it is recognized as a meandering and often discontinuous sulcus called the intraparietal sulcus that tends to run in a parasagittal plane. We will return to the superior parietal lobule when we explore the medial surface of the hemisphere. The gyral formations just inferior to the intraparietal sulcus include the supramarginal gyrus and the angular gyrus, two principal components of the inferior parietal lobule. The supramarginal gyrus usually forms a “horseshoe” shape around the posterior limit of the lateral fissure, with the angular gyrus just posterior to the supramarginal gyrus. Like the shape of the lateral fissure itself, these gyral formations can be quite different between the two hemispheres; so do find these inferior parietal gyri in both hemispheres of the same brain. Recall the somewhat arbitrary means for defining the lateral boundary that marks the division of the posterior hemisphere into the parietal, temporal and occipital lobes. This lateral boundary is an imaginary line between the parieto-occipital sulcus dorsally and the pre-occipital notch ventrally (see Figure 1.9B). Simply put, all gyral structures posterior to this line are occipital and have some role to play in elaborating visual perception. Fortunately for you, the complexity and inter-individual variation of these gyri defies easy application of standard nomenclature; for this reason, we will refer to these gyral structures as lateral occipital gyri and leave the business of naming these gyri for the cortical cartographers.Refer to the blank figure below, which is a photograph of the lateral surface of a human brain and see if you can name all of the bolded features above. Feel free to label and annotate this hard copy as you become more familiar with recognizing the names and locations of the important gyri, sulci and fissures of the lateral hemisphere.
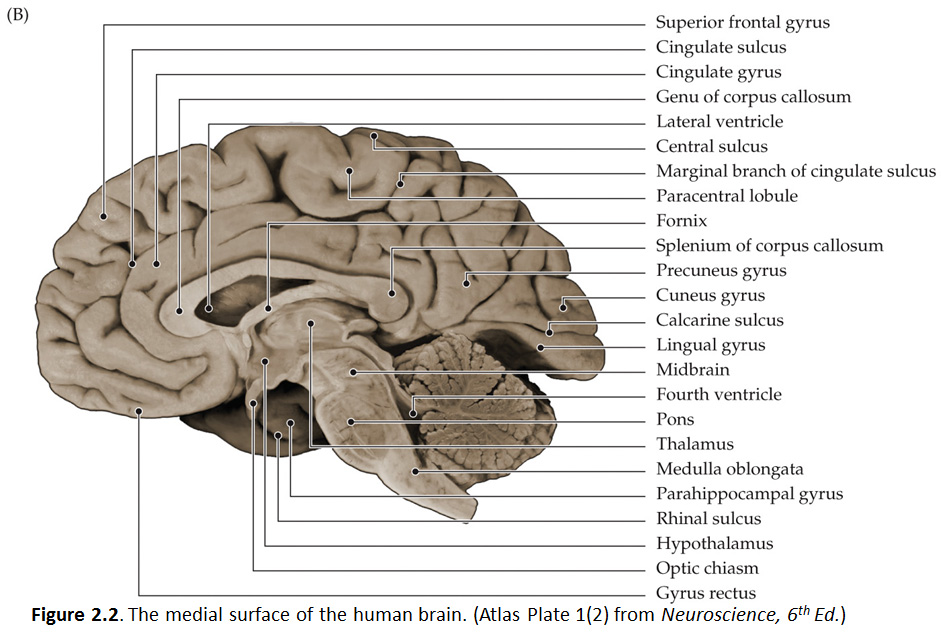
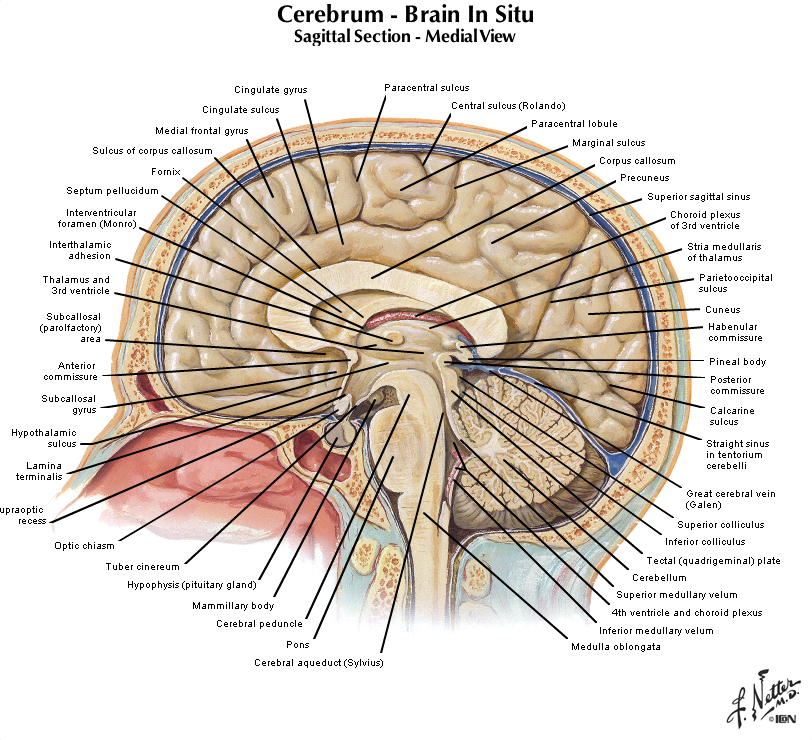
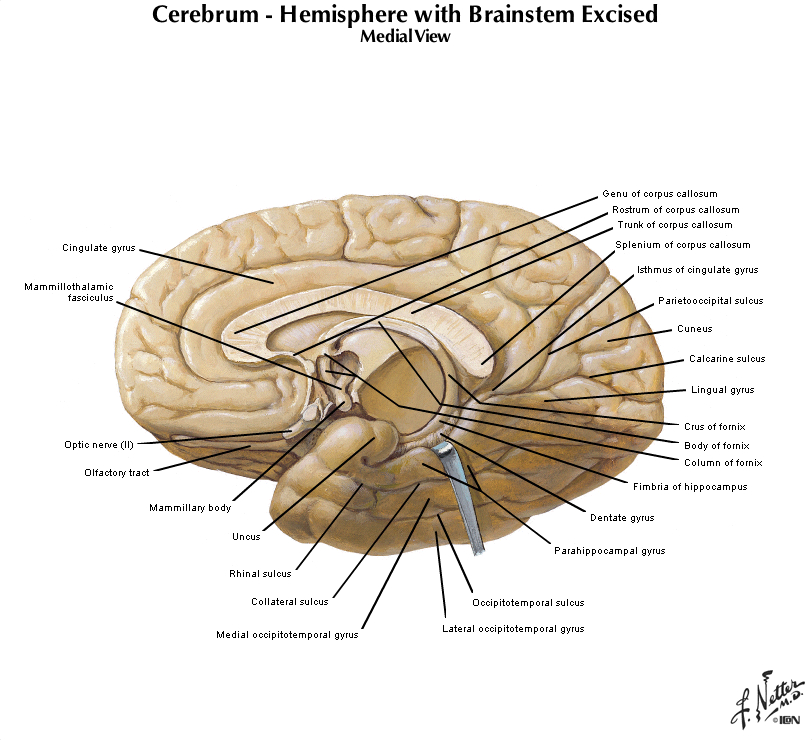
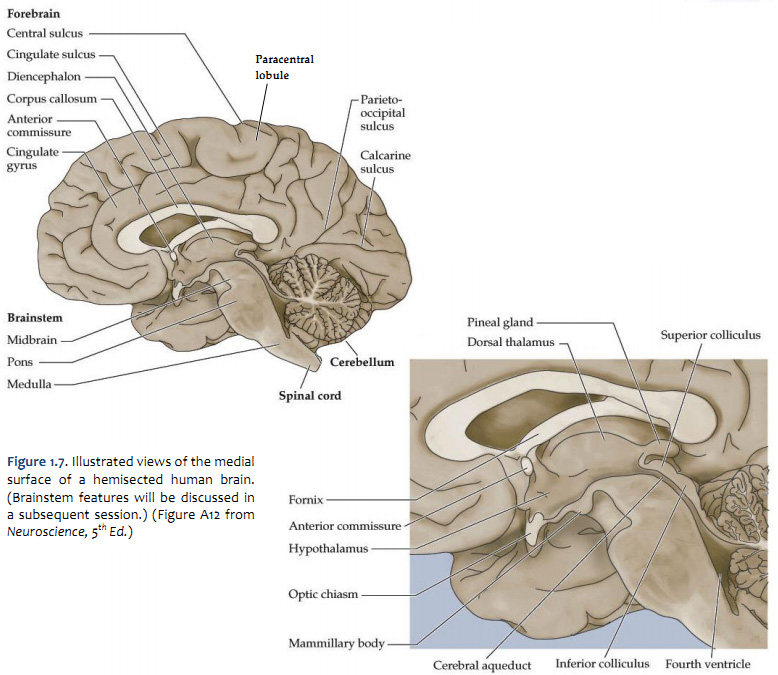
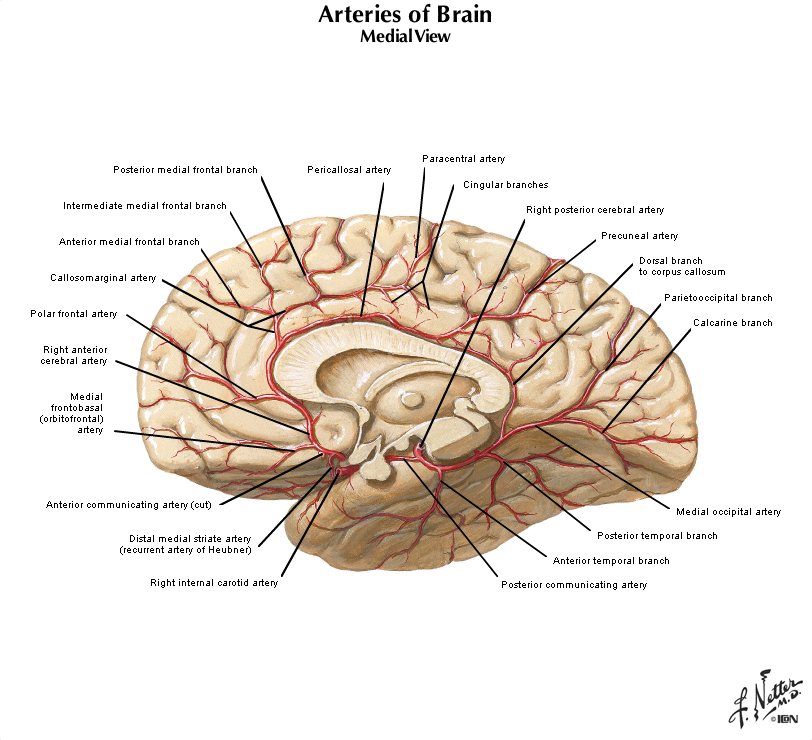
Let's turn our attention now to the mid-sagittal plane, so get in front of you a hemisphere from a brain sectioned in that plane or a brain model with the two hemispheres separated and refer to Figure 2.2 (see also Grants 8-87a, Netters 100A, and Netters 110B). Beginning from the superior margin of the hemisphere, the most anterior and dorsal gyral formation is simply the medial continuation of the superior frontal gyrus. A long, almost horizontal sulcus, the cingulate sulcus, extends across the medial surface of the frontal and parietal lobes just below the superior frontal gyrus (if you performed the Considering the Central Sulcus self-study exercise, then you are already familiar with the course of cingulate sulcus). The prominent gyrus below the cingulate sulcus is, of course, the cingulate gyrus. Along with the cortex adjacent to the cingulate gyrus, this gyral formation wraps around the corpus callosum and lateral ventricle into the temporal lobe. This extended rim (Latin, limbus) of cortex is sometimes called the 'limbic lobe'. These cortical areas—and the subcortical areas connected to them, together with additional telencephalic structures in the temporal lobe and ventral frontal lobe—are often referred to as the 'limbic system'. However, this so-called system (so-called primarily for historical reasons) is not unimodal, as the term 'system' implies. Rather, the limbic 'system' is involved in the regulation of visceral motor activity, emotional experience and expression, olfaction, and memory, to name some of its better understood functions (see Appendix 4 for more information on the anatomy of the limbic forebrain).
The caudal portion of the superior frontal gyrus forms the paracentral lobule, as it joins the medial continuation of the pre- and post-central gyri. Just as on the lateral surface of the hemisphere, on the medial face of the hemisphere the frontal lobe extends from the central sulcus forward. At the inferior margin of the frontal lobe is the medial aspect of an inferior gyrus called the gyrus rectus (see 'ventral view' below), and a small division of the anterior and inferior cingulate gyrus, called the subcallosal (subgenual) area, which is just below the genu ("knee") of the corpus callosum. This subcallosal area has become an important target for deep brain stimulation in the treatment of various psychiatric diagnoses, including major depressive disorder. Locate again the paracentral lobule and note that its posterior bank forms the marginal (dorsal) branch of the cingulate sulcus. Between this marginal branch of the cingulate sulcus and the parieto-occipital sulcus (which as you will recall divides the parietal and occipital lobes) is the precuneus gyrus. The name of the gyrus on the medial face of the parietal lobe will make sense as you read on. The precuneus gyrus may be understood anatomically as the medial continuation of the superior parietal lobule, which we otherwise recognize on the dorsal-lateral surface of the hemisphere. Next, reorient yourself to the parieto-occipital sulcus (see Figure 1.13) and find the calcarine sulcus, which intersects the parieto-occipital sulcus at nearly a right angle and extends typically to the occipital pole of the hemisphere (search this term in the Sylvius4 Online Visual Glossary and click on the loudspeaker icon to hear the proper pronunciation of the calcarine sulcus; note that this sulcus is particularly straight in the Sylvius brain—it is often more curvilinear). We'll come back to this part of the brain when we study the visual system. For now, notice the "tongue"-like gyral structure that forms the inferior bank of the calcarine sulcus, and the "wedge"-shaped gyrus that forms its superior bank. Thankfully, the formal terms for these gyri mean just that:
The precuneus gyrus, of course, lies just in front of (anterior to) the cuneus gyrus, and now you know why it was given this name (the gyrus in front of the "wedge").
Refer to the figure below, which is a photograph of the medial surface of a human brain and see if you can name all of the bolded features above. Feel free to label and annotate this hard copy as you become more familiar with recognizing the names and locations of the important gyri, sulci and other visible features of the medial hemisphere in mid-sagittal plane.
To further consolidate what you have learned about the convolutions of the cerebral hemispheres, try performing Challenge 2.1 (below) as one suggested means for conceptualizing your understanding of these primary gyral formations.
Although the major objectives of this portion of the lab session are centered around the features of the lobes of the cerebral hemisphere, we should take advantage of the midsagittal hemispheres that are available and introduce a few more features of the brain that are visible in these specimens. Let’s begin with three prominent fiber bundles (i.e., bundles of axons extending from one part of the brain to another) associated with the cerebral hemispheres that can be seen from the medial view (see Figure 1.12). These are:
In the view shown in Figure 1.12, the axons in the corpus callosum and anterior commissure are running perpendicular to the plane of the page, and the visible fibers of the fornix are running within the plane of the page. The other subdivisions of the brain, all of which can be seen in Figures 1.10 and 1.12, are as follows:
All components of the ventricular system, except perhaps for the lateral ventricles, can be seen on a typical medial surface of the brain cut in the midsagittal plane. In Figure 1.12, the lateral ventricle is visible in this hemisphere because the septum pellucidum has been dissected away; this is a very thin structure made of ependymal cells that forms the medial wall separating the two lateral ventricles. The third ventricle forms a narrow space in the midline region of the diencephalon, between the one that you see and the one that has been cut away. The communication of the third ventricle with the lateral ventricle is through a small hole, the interventricular foramen (or foramen of Monro), at the anterior-dorsal end of the third ventricle just lateral to the fornix. The third ventricle is continuous caudally with the cerebral aqueduct, which runs through the midbrain. At its caudal end, it joins the fourth ventricle, a large space in the dorsal pons and medulla. The fourth ventricle narrows laterally and caudally where cerebrospinal fluid (CSF) exists the adult ventricular system via two lateral apertures (the foramen of Luschka) and a single medial aperture (the foramen of Magendie). CSF then circulates throughout the subarachnoid space before returning to the venous circulation in the superior sagittal sinus.
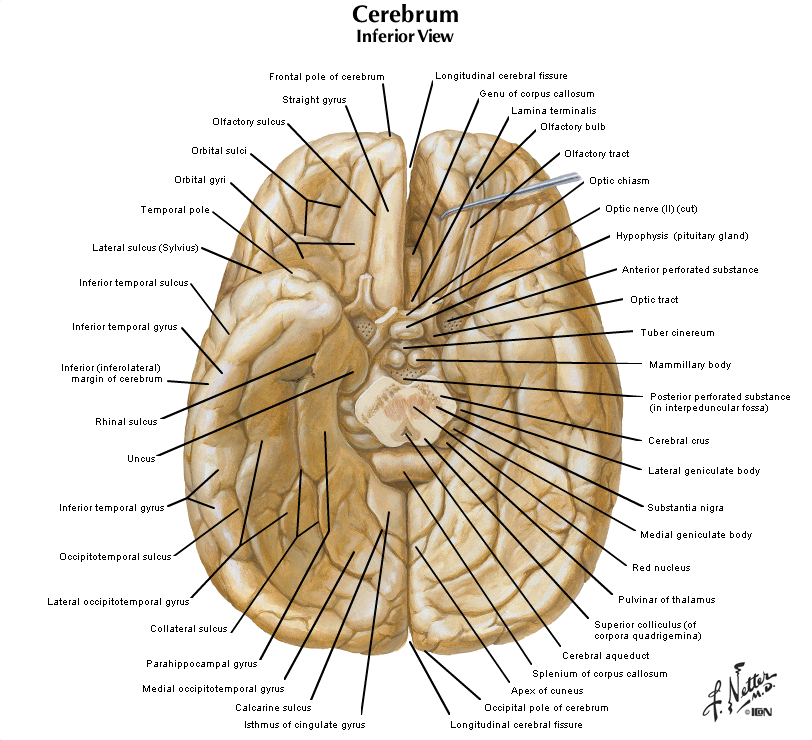
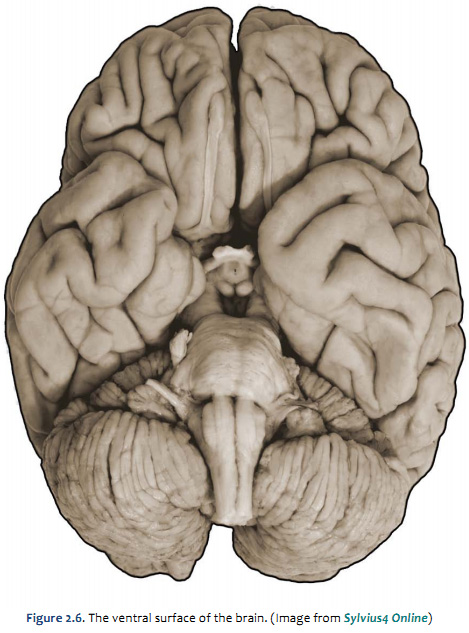
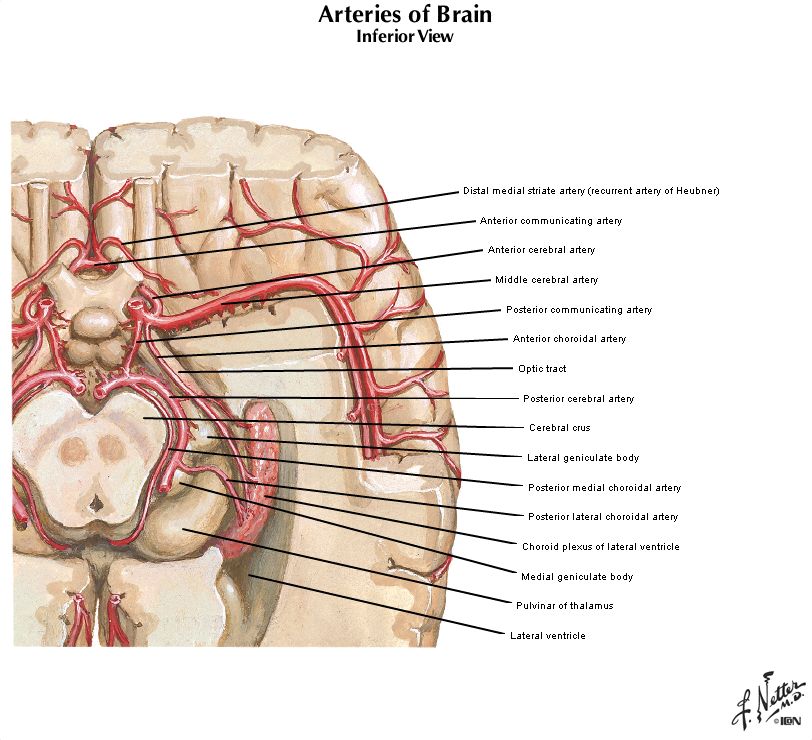
Figure 2.4 (see also Netter 101) provide another look at the surface of the whole brain. Most of the subdivisions of the brain can be seen when it is viewed from its ventral aspect. The inferior surfaces of the frontal and temporal lobes of the cerebral hemispheres are prominent in this view. Running along the inferior surface of the frontal lobe near the midline are the olfactory tracts, which arise in little swellings at their anterior ends, the olfactory bulbs. The olfactory bulbs receive the axons of sensory cells in the olfactory mucosa (these axons are the first cranial nerve), and neurons in the bulbs give rise to fibers in the olfactory tracts (therefore, the tracts are part of the forebrain). Just superior to the bulbs, of course, is the ventral aspect of the frontal lobe, often referred to as the "orbital cortex" since this is the portion of the frontal lobe the overlies the orbits. The olfactory bulbs and tracts lie in the olfactory sulci (one in each hemisphere). This sulcus divides the gyrus rectus at the medial margin of the ventral frontal lobe from the more complex gyral structures that occupy much of the remaining ventral aspect; we will refer to these gyri simply as orbital gyri. Most of the ventral aspect of the frontal lobe is visible in Figure 2.4 (see also Netter 101) except for that posterior portion hidden by the underlying anterior temporal lobes. On the ventral surface of the temporal lobe, the inferior temporal gyrus occupies most of the visible surface of the lobe (with brainstem and cerebellum intact). However, it is possible to appreciate additional gyral structures on the medial side of the ventral temporal lobe. The medial boundary of the inferior temporal gyrus is formed by two sulci, the rhinal sulcus more anteriorly and the collateral sulcus more posteriorly. Just on the medial side of these sulci are gyral structures that are associated with the hippocampal formation (a primitive cortical structure that we will see when we dissect the brain); first, is the parahippocampal gyrus and then a medial protuberance of this gyrus called the uncus. Just posterior to the parahippocampal gyrus is another prominent structure called the occipito-temporal gyrus (also called—especially by functional brain imagers—the fusiform gyrus), but this gyrus is mostly hidden from view in Figure 2.4 by the cerebellum (and therefore not labeled). A small part of the diencephalon is visible in this view of the brain. The part that you see is the hypothalamus, bounded rostrally by the optic chiasm (formed by the crossing of some of the axons in optic nerves) and caudally by the mammillary bodies, which are considered part of the hypothalamus. The midbrain is mostly hidden from view by the temporal lobes; however, the prominent paired cerebral peduncles are visible (these structures define a space between them called the interpeduncular fossa). The pons is obvious in this view, as is the cerebellum. Caudal to the pons is the medulla oblongata. The columnar swellings on its ventral surface on either side of the midline are known as the medullary pyramids. They contain axons that arise mainly in the precentral gyrus (the motor cortex) and terminate in the spinal cord (i.e., the corticospinal tract) and medulla (a portion of the corticobulbar tract). Lateral to the pyramids are the inferior olives. Caudal to the medulla, a portion of the cervical spinal cord is seen. We will consider the brainstem in much more detail in later sessions, when we identify the cranial nerves and study their relation to external and internal features of the brainstem. For now, this brief survey of the ventral surface of the brain (including the brainstem) will facilitate understanding the distribution of major arteries that convey blood supply to the brain—a topic that we now turn to in the second half of this laboratory experience. Refer to Figure 2.6 below, which is a photograph of the ventral surface of a human brain, and see if you can name all of the bolded features above. Feel free to label and annotate this hard copy as you become more familiar with recognizing the names and locations of the important gyri, sulci and other visible features of the ventral surface.
The remainder of this lab will challenge you to focus on the blood supply to each embryological subdivision of the brain, including the cerebral hemispheres and the brainstem. For completeness, we will also briefly review the blood supply to the spinal cord. Lesions involving the different vessels and their branches lead to specific syndromes that are easy to understand if you learn the distribution patterns of the vessels and the organization of the brain and spinal cord subdivisions that they supply.
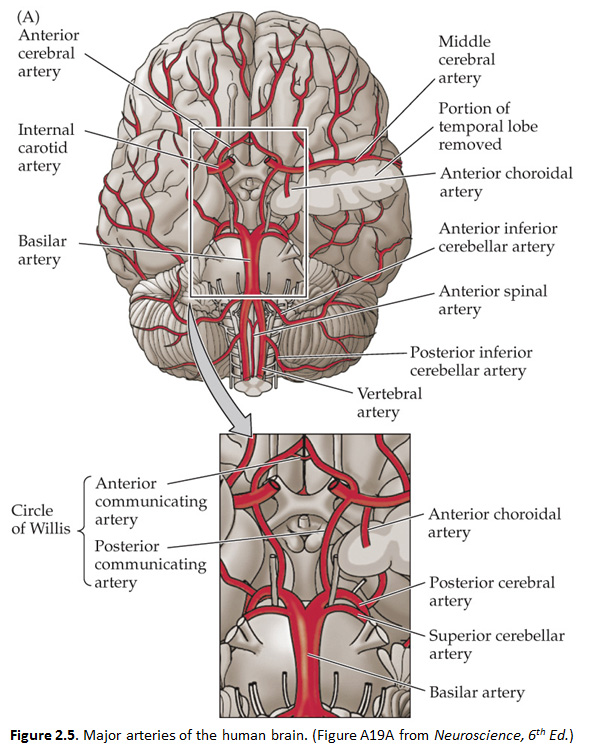
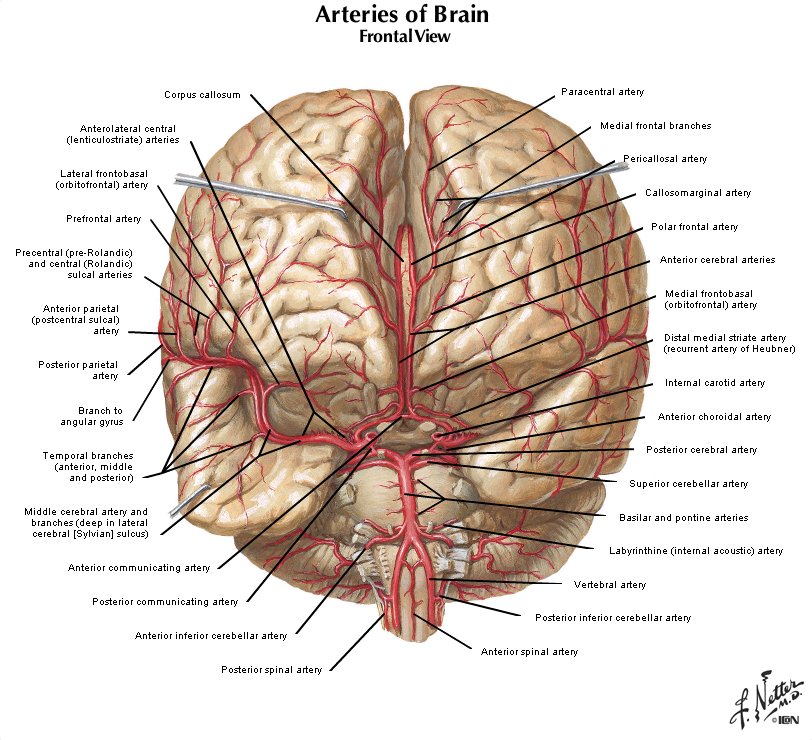
As you learned in Normal Body, the brain receives its arterial supply from two sources: the internal carotid arteries and the vertebral/basilar arteries (the two vertebral arteries join to form the basilar artery at the base of the pons). Both the internal carotid arteries and the vertebral/basilar arteries give rise to four main branches, commonly referred to as the anterior circulation and the posterior circulation, as depicted in the chart below and in Figure 2.5. This chart accounts for the major branches of the carotid and vertebral/basilar arteries. The two systems of arteries are joined at the junction between the posterior communicating artery and the posterior cerebral artery. In most humans, the posterior cerebral artery receives its blood supply from the vertebral/basilar system. In some people, the posterior communicating artery is quite large, and the posterior cerebral artery may be perfused by the carotid artery. Generally speaking, the anterior circulation supplies the forebrain (the cerebral hemispheres and the diencephalon), and the posterior circulation supplies the brainstem and the upper spinal cord. However, for most people, the arterial supply to the CNS is not quite that simple. As just mentioned, the posterior cerebral artery supplies the posterior forebrain, including some deep structures, and it also supplies parts of the midbrain in the brainstem. Thus, as indicated in the chart (by listing the posterior cerebral artery twice, once in each group), the posterior cerebral artery contributes to both the anterior and posterior circulations. As you study this chapter, you should learn the distributions of these 10 arteries listed in this chart.
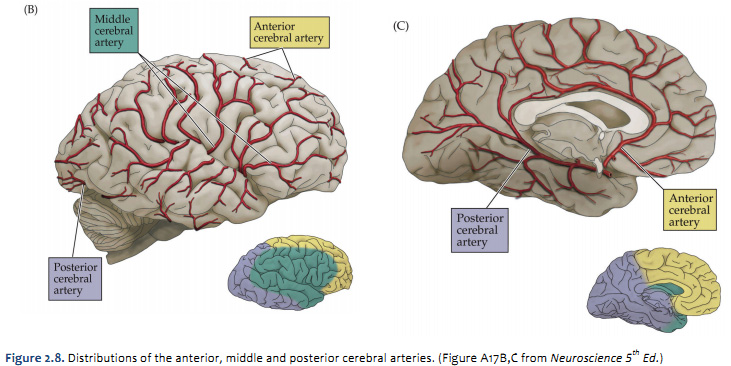
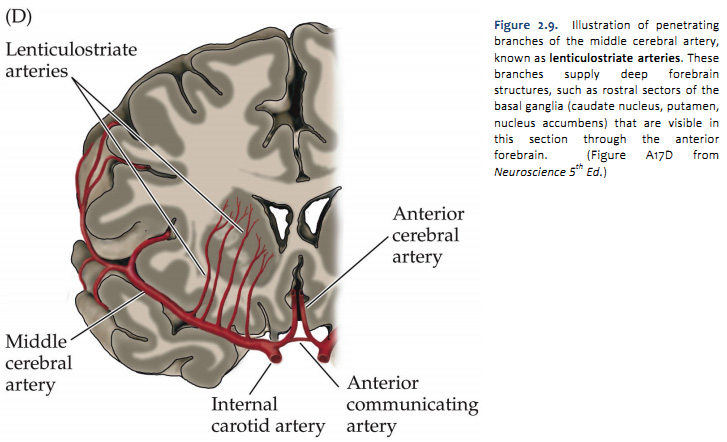
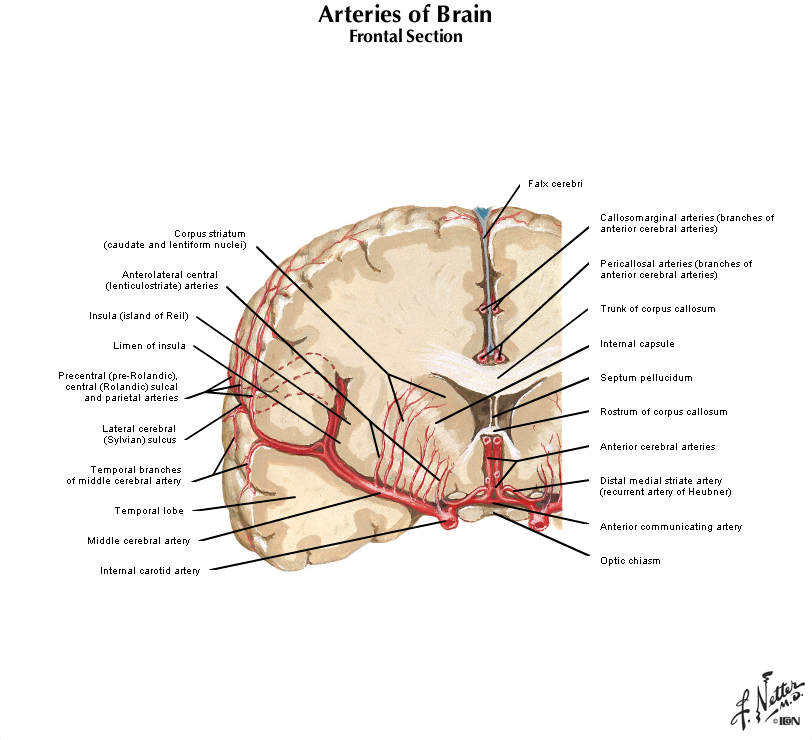
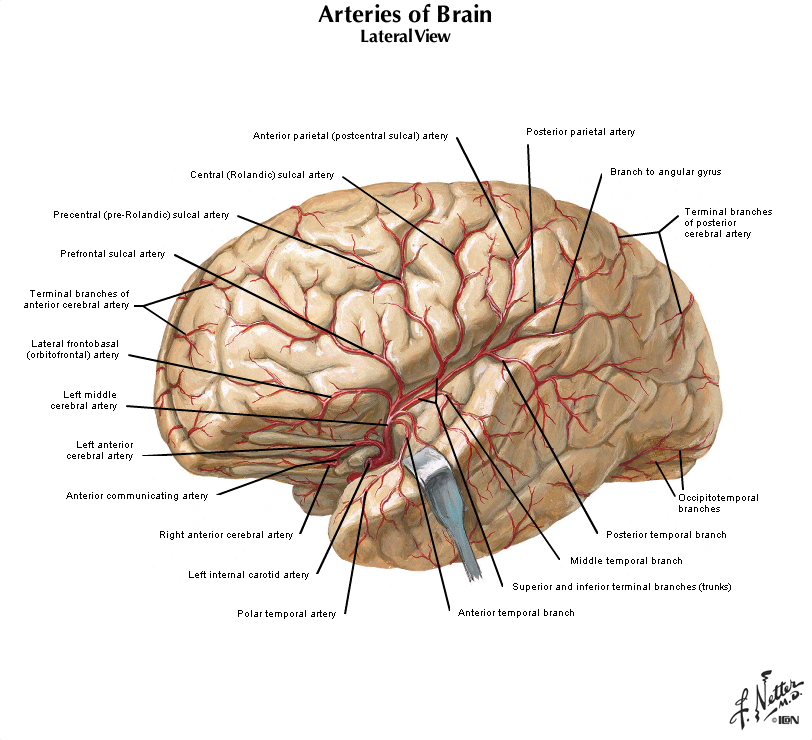
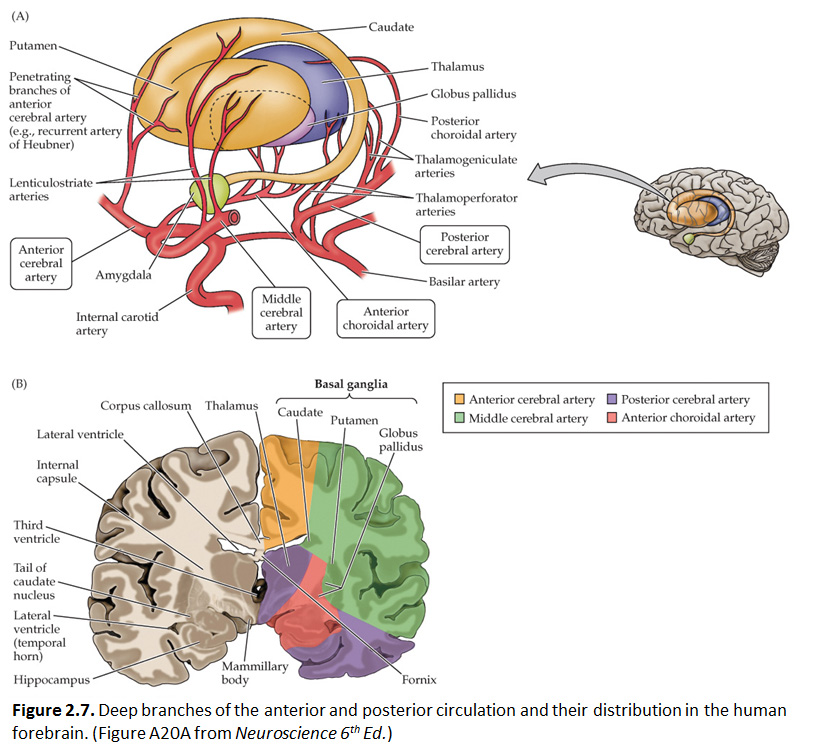
The four major arteries that arise from the internal carotid artery plus the posterior cerebral artery form the anterior circulation. The pattern of branching of each artery is similar: each gives rise to branches that supply cortical structures and each gives rise to branches that penetrate the ventral surface of the brain and supply deep structures (the basal ganglia, thalamus and internal capsule), as illustrated in Figures 2.8 & 2.9. (see also Netter 134A, 134B, 135A, and 135B). The branches that supply deep structures are known collectively as perforating arteries, central arteries, striate arteries, or ganglionic arteries.) An extensive region of the central and lateral cerebral hemispheres is supplied by the middle cerebral artery (green shade in Figure 2.8, see also Netter 135A). Included in this region are the sensorimotor areas that govern the upper extremities and face, and the language areas of the left hemisphere (Broca’s area and Wernicke’s area). The anterior cerebral artery supplies regions in the medial aspect and dorsal and orbital margins of the frontal lobe, and the medial aspect and dorsal margin of the anterior parietal lobe (yellow shade in Figure 2.8, see also Netter 135B). Included in this extended territory are sensorimotor areas in the paracentral lobule that govern the lower extremity, accessory motor areas in the cingulate gyrus that govern the upper face (see Clinical Applications Box in Chapter 17 of Neuroscience, 6th Ed.), and limbic areas in the medial frontal lobe. The posterior cerebral artery supplies regions in the posterior parietal lobe, inferior temporal lobe and occipital lobe (blue shade in Figure 2.8, see also Netter 135B). Included in this region are primary and associational (higher-order) visual areas in each lobe and ‘limbic’ regions in the posterior cingulate and parahippocampal gyri. At this point, you should recognize that this chapter on blood supply to the brain (and spinal cord) affords the opportunity to review what you have already learned regarding the localization of function in the cerebral cortex [Challenge 2.2] (below).
Each of the four major branches of the internal carotid artery give rise to penetrating branches, in addition to the superficial branches just described, that supply gray and white matter structures deeper in each hemisphere. These deep branches follow a reasonably straightforward, anterior-to-posterior pattern of branching (Figure 2.10). The anterior cerebral artery supplies the anterior caudate and putamen, the nucleus accumbens, and the anterior limb of the internal capsule. The middle cerebral artery supplies the body of the caudate and most of the putamen, most of the globus pallidus, the middle part (or genu) of the internal capsule, and the anterior hypothalamus. These deep penetrating branches of the middle cerebral artery are usually called the lenticulostriate arteries (see Figure 2.9; see also Netter 134B). The anterior choroidal artery (Netter 132B) supplies the amygdala, hippocampus, the anterior part of the thalamus, part of the globus pallidus, and the choroid plexus of the lateral ventricle. The posterior communicating and posterior cerebral arteries supply the posterior hypothalamus, most of the thalamus, and the choroid plexus of the third ventricle. (Branches of the posterior cerebral artery also supply the midbrain as described later.) It is not necessary to remember all of these details! However, it is easy to remember that the deep structures of the forebrain are divided approximately into four sectors progressing from anterior to posterior, and a different artery perfuses each sector (Figure 2.10).
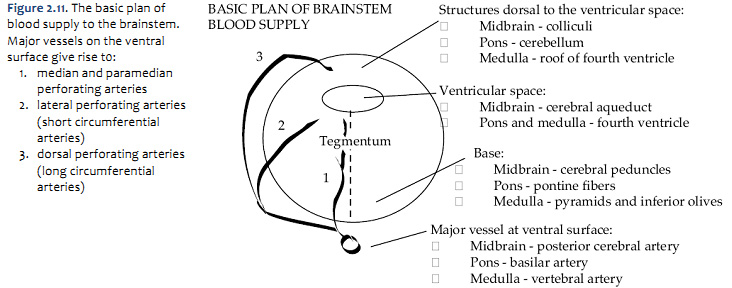
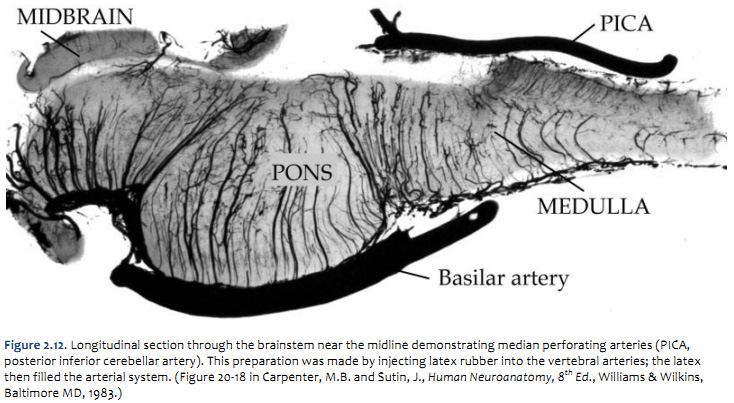
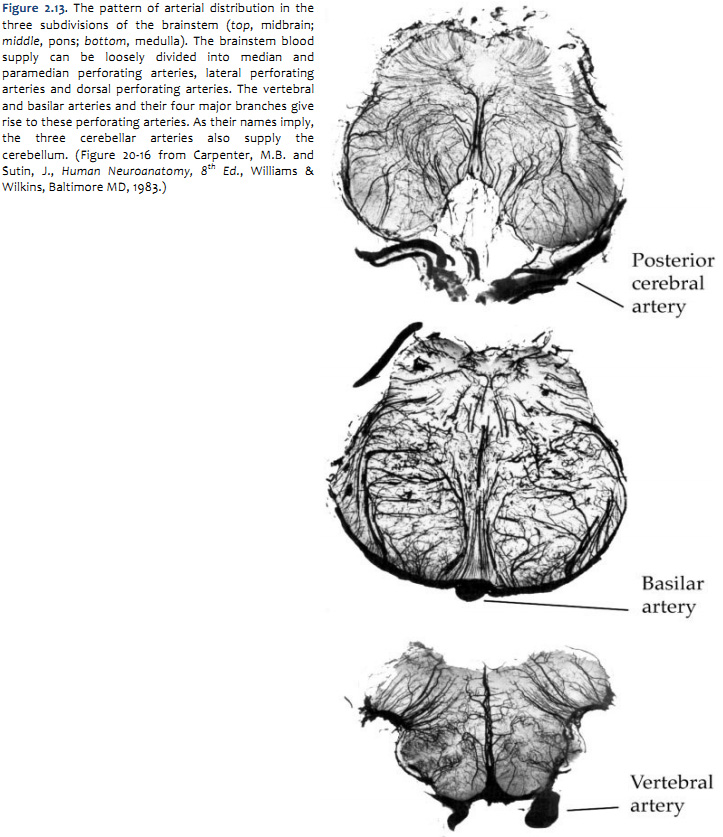
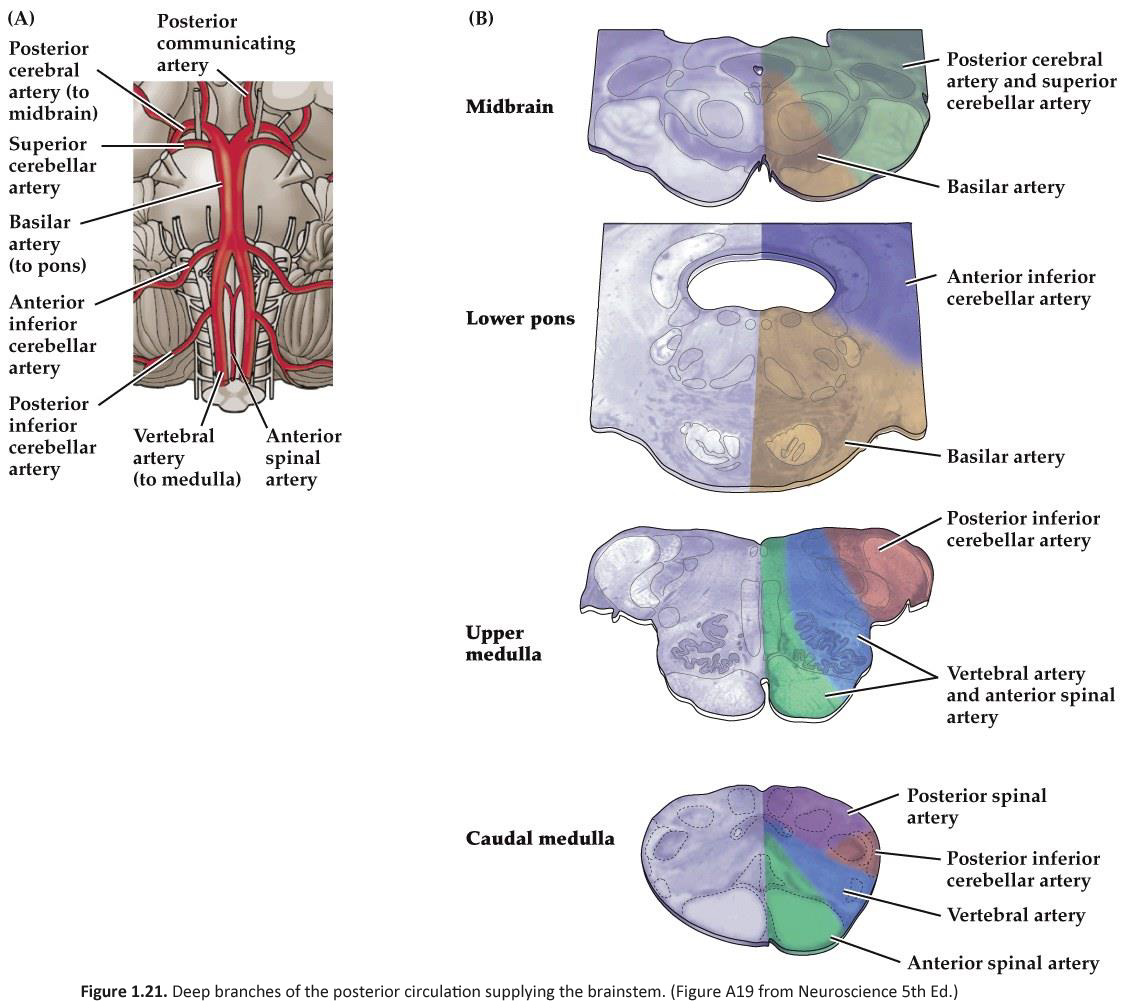
Arterial supply of the brainstem
The pattern of arterial distribution is similar in all three subdivisions of the brainstem:
As you study Figure 1.21, refer frequently to the inset in the upper left, which shows an illustration of the posterior circulation as viewed from the ventral aspect of the hindbrain. Recognize how the major vessels and their circumferential branches are related to the divisions of the brainstem. Also, note that the cerebellar arteries give rise to penetrating branches that supply the more lateral and dorsal aspects of the brainstem, as they course toward their respective regions of the cerebellum (superior cerebellum, anterior-inferior cerebellum and posterior-inferior cerebellum). Similarly, In the caudal subdivisions of the medulla oblongata, the spinal arteries (not named in the chart above) contribute to the supply of blood in medial and dorsal-lateral sectors of the junctional region between the brainstem and the spinal cord (see lower panel in Figure 1.21B). Thus, each of the three subdivisions of the brainstem can be divided into medial and lateral ‘wedges’ of tissue that are supplied by different perforating branches. Vascular lesions that affect individual wedges of brainstem tissue lead to distinct neurological syndromes, as you will study later in this course in the context of understanding the organization of long pathways in the brainstem and the distribution of cranial nerve nuclei. For now, note that most vascular lesions of the brainstem are usually unilateral, since each side of the brainstem is supplied by different sets of circumferential vessels. However, bilateral damage may result if the basilar artery itself is blocked, since it gives rise to vessels that supply both sides of the brainstem. As you will discover below, similar bilateral lesions will result with blockage or hemorrhage of the anterior spinal artery, since it too gives rise to branches that supply both sides of the ventral spinal cord. Drainage of Venous Blood
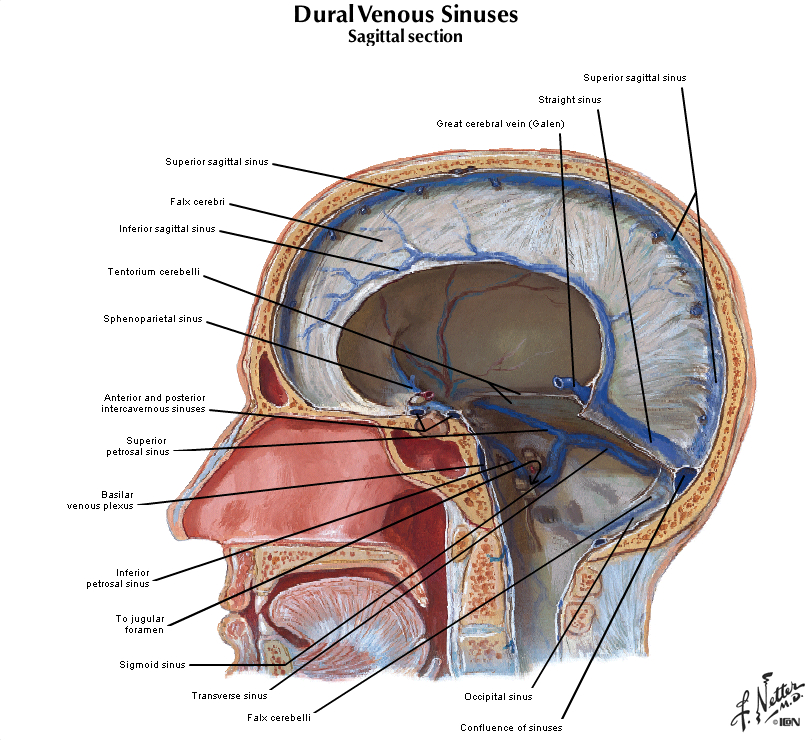
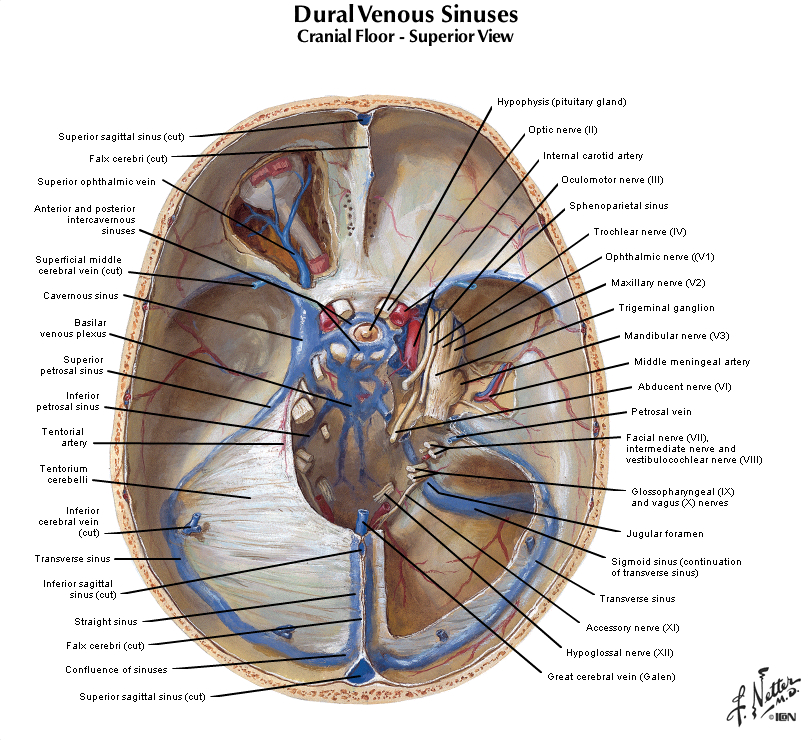
Figure 2.14 (see also Netter 97 and 98A) illustrates in the system of veins that provides for the drainage of venous blood from the brain and cranium. This figure provides an overview of the means by which blood completes its passage through the brain from the arterial vasculature back to the heart via the internal jugular veins. In brief, the more superificial veins of the cerebrum drain into the superior sagittal sinus along the dorsal midline of the hemisphere, or the cavernous sinus in the base of the cranium. The deeper veins of the brain drain into the inferior sagittal sinus at the inferior margin of the falx cerebri, and the great vein of Galen, which in turn join to form the straight sinus. The major venous sinuses inside the cranium are formed by a separation of the two layers of dura, which should be appreciated by inspecting samples of dura that are available in the laboratory. The superior sagittal sinus and the straight sinus drain into a pair of transverse sinuses, which are oriented roughly in the horizontal plane along the posterior margin of the tentorium. The transverse sinuses then turn in the inferior direction, becoming the sigmoid sinuses, which finally exit the cranial vault as the internal jugular veins. Recall that in addition to providing venous drainage for the brain and meninges, the venous sinuses also serve as a resorption site for cerebrospinal fluid at arachnoid granulations (illustrated in Netter 103)
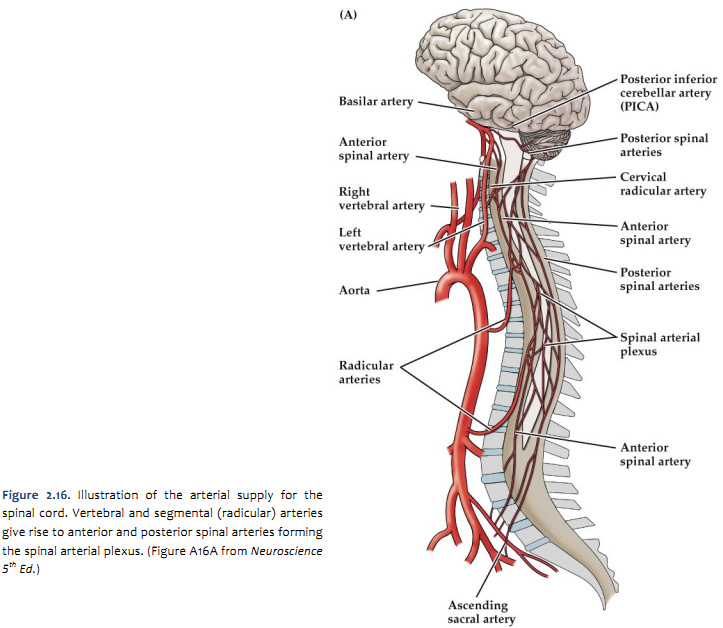
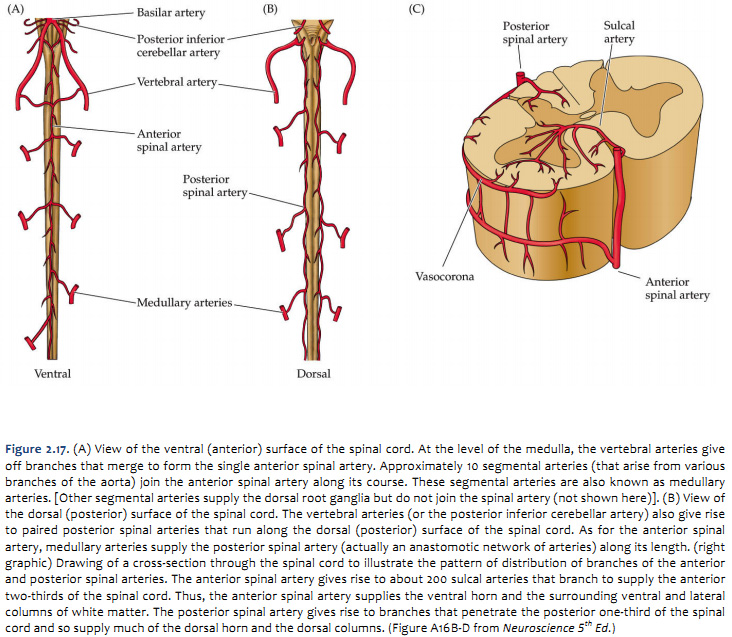
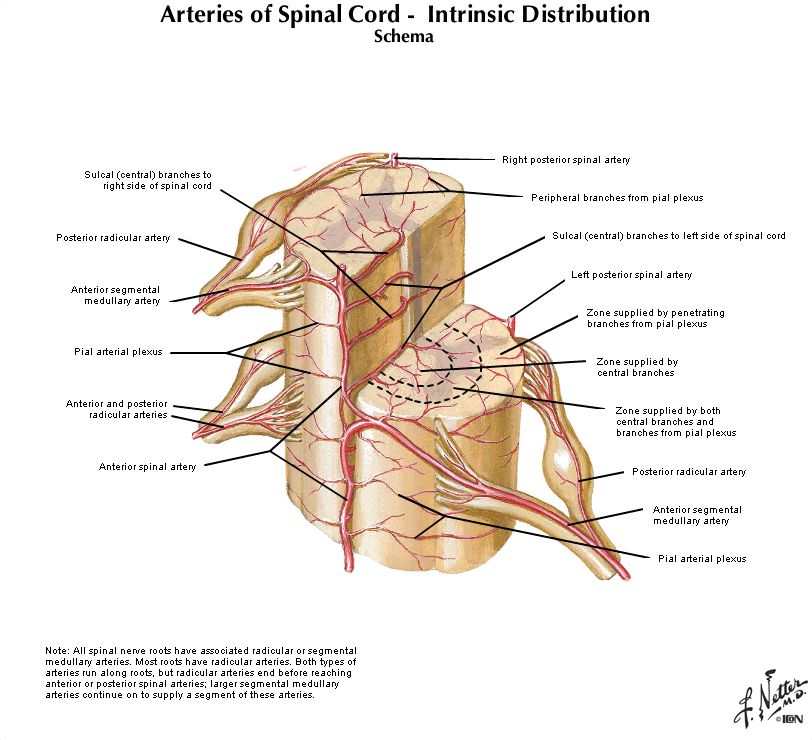
The arterial blood that supplies the spinal cord comes from two sources: the vertebral arteries and segmental arteries that arise from branches of the aorta. These arteries join the anterior and posterior spinal arteries as illustrated in Figure 2.16 and 2.17 (see also Netter 164, 165A, 165B). It is important to realize that if any of the medullary arteries are obstructed or damaged, blood supply to part of the spinal cord may be compromised and neurological damage will result. The pattern of damage depends on whether the supply to the posterior arteries or anterior artery is interrupted. An anastomotic network of vessels known as the vasocorona connects these two sources of supply and sends branches into a narrow zone of white matter around the margin of the spinal cord (see Figure 2.17 and Netter 165B). The vasocorona may be sufficient to supply the most lateral white matter in cases in which the anterior spinal artery is occluded. At the level of the medulla, the vertebral arteries give off branches that merge to form the single anterior spinal artery. Approximately 10 segmental arteries (that arise from various branches of the aorta) join the anterior spinal artery along its course. These segmental arteries are also known as medullary arteries. [Other segmental arteries supply the dorsal root ganglia but do not join the spinal artery (not shown here)]. The vertebral arteries (or the posterior inferior cerebellar artery) also give rise to paired posterior spinal arteries that run along the dorsal (posterior) surface of the spinal cord. As for the anterior spinal artery, medullary arteries supply the posterior spinal artery (actually an anastomotic network of arteries) along its length. The posterior spinal arteries give rise to branches that penetrate the posterior one-third of the spinal cord and so supply much of the dorsal horn and the dorsal columns. The anterior spinal artery gives rise to about 200 sulcal arteries that branch to supply the anterior two-thirds of the spinal cord. Typically, unilateral sulcal arteries alternate with one supplying one side of the midline followed within a few millimeters by another supplying the opposite side. Thus, the anterior spinal artery supplies the ventral horn and the surrounding ventral and lateral columns of white matter.
|
|
Click here to submit questions or comments about this site. Duke University | Duke Medicine | School of Medicine | School of Nursing | Doctor of Physical Therapy Updated 1/02/18- Velkey |
||